Ctarenie v celičnih kulturah
Video: Jeunesse Globalni FINITI ™ Mobilne senescenco konča tukaj
Vsebina
Še posebej, metoda celične kulture omogoča reševanje te pomembne naloge.
1. Primerjava kulture celic, ki izvirajo od dajalcev, različnih starosti, je mogoče razlikovati med spremembami, povezanih s staranjem sestavnih inducirane. Na primer, je bilo ugotovljeno, da je mitotični aktivnosti človeških fibroblastnih kultur, ki so pod enakimi pogoji, pada z naraščajočo starostjo organizma donatorskih polj (Schneider, Mitsui, 1976- Schneider, 1979).
Zato opazili senescentnih tkivih organizmi zmanjšujejo mitotićnega dejavnosti (Gelfant, Smith, 1972- Cameron, Thrasher, 1976), ki je povezana s sestavnimi celic sprememb samih.
Po drugi strani pa povečanje, povezanih s starostjo, v tirozin indukcijsko obdobje jetra pod vplivom hidrokortizona (Adelman, 1972), je treba upoštevati inducirane zaradi različnih starosti v kulturah hepatocitih podgan pričakujemo nobenih razlik med indukcijo encima (Britton s sod., 1976). Tako je aplikacija omogoča celičnih kultur za določitev narave staranja.
2. Postopek za celične kulture ponuja edinstveno priložnost za raziskovanje in natančno študijo o konstitutivni biokemične (Vilenchik et al., 1979) in citologija (Schneider, 1979), sprememb, povezanih s starostjo. Ta metoda omogoča tudi neposredno preverjanje teorije staranja (Danner et al., 1978).
3. Primerjalna študija celičnih kultur, pridobljenih iz različnih živali, omogoča, da se opredeli značilnosti celic posebnega pomena za biologiji staranja. Na primer, je bilo ugotovljeno, da so bile živali pozitivno korelira z prostate fibroblastov v kulturi sposobnost za popravilo UV povzroča poškodbe DNK (Hart, Setlow, 1974) in v negativni korelaciji z intenzivnostjo vezavo rakotvornih snovi na DNK fibroblastov teh živali (Schwartz, Moore, 1977, 1979). Zato, mehanizmi za zaščito genoma pred poškodbami so za gerontologijo poseben interes.
4. Dolgoročno kultura (približno eno leto) in vitro celice lahko model številne spremembe, ki se pojavljajo med staranjem. Čeprav je vprašanje o ustreznosti vitro POGOJI vivo ostaja odprto vprašanje, staranju celične kulture je namenjen za povečanje števila preskusnega pregleda (Hay, 1967- Bershad, Gelfand, 1970- Cristofalo, 1972- Hayfiick, leta 1977 Gavrilov, Gavrilova, 1978) in teoretično delo (Olovnikov, od leta 1973 Holliday, 1975 Hirsch, od leta 1978 Gavrilov, Yaguzhinskii, 1978).
Merila in razloge za smrt celičnih kultur
Trenutno nameščeni Limited pričakovana življenjska doba (RV) naslednje celične kulture normalnih Človeške fibroblastne celice (Hayfiick, Moorhead, 1961- Hayfiick, 1965- Harper, Grove, 1979), konji, več vrst kenguruji (Stanley et al, 1975), marsupials Potorous tridactylus (Cristofalo, 1972- Stanley et al, 1975), želve (Goldstein, 1974), kun (Hayfiick, 1975), piščančja (Kaji, Matsuo, 1979) in miš (Hayflick, 1975, 1977) -. epidermalne keratinocite (Rheinwald, Green, 1977), celica glia (Brunk`et al., 1973) in humane jetrne (Kahn in sod., 1977a, 1977b) običajnih hondrocitov (Mayne s sod., 1976) in Rous sarkoma virusom transformirana chick fibroblasti (Ponten, 1970- Leblond-Larouche, Morais , 1976).Vendar pa je kritična analiza podatkov iz literature (Gavrilov, Gavrilova, 1978), se zdi, da še vedno ni splošno sprejete kvantitativna merila za celične kulture smrti, in sami koncepti - "smrt" in "življenjska doba" kulture - zelo pogojno in subjektivne.
Večina raziskovalcev se imenuje kultura mrtva, če v poljubno vnaprej določenem časovnem intervalu (običajno en do štiri tedne) preneha povečati svojo moč na želeno vrednost, kljub ugodnim pogojem za rast (običajno določi število, 2-4 krat število začetne kulture) .
Po tej definiciji je smrt celic v kulturi ni potrebno, kot vsako kulturo sposobnih, vendar se ne delijo celice, v tej opredelitvi, se šteje mrtev. Poleg tega je kultura mrtvih še naprej počasi narašča, čeprav imajo nizko mitotićno aktivnost in vsebujejo samo 10-20% celic, ki lahko replikacije sinteze DNA (Cristofalo, Sharf, leta 1973 Milo, Hart, 1976).
Takšne kulture so včasih spontano obnoviti normalno rast in nato nekaj podvojitev prebivalstva pred naslednjo "smrtno" (Holliday, Tarrant, 1972). Masa celične smrti ob koncu kulture življenja, ki je priložen velik pomen zgodnjih delih (Hayfiick, Moorhead, 1961- Hayfiick, 1965), je bil artefakt posevki operacijo.
Dejstvo, da z gojenjem celice redno kultivirajo, t. E. Pritrjena na predelavo substrat celične plasti 0,1% raztopine tripsina (tripsinizacija) in ločevanja celic odsidranje enojno jo s hidrodinamične učinke (obešanje).
V prvih die velikih celic (milo, 1973), v katerem je večina starih in mrtvih kulture (Bowman et al., 1975 Mitsui, Schneider, 1976). Poleg tega je v teh kulturah povečalo število celic stikov, ki jih ni mogoče uničena med subkulture, ne da bi pri tem poškodovali celice same (milo, 1973).
Zato je v razmerah nizke rasti kulture pri konstantni uničenje celic v procesu ponovno zasaditev vtis popolne lizi. V tistih primerih, v katerih so bile izključene vpliv posevki poslovanja, niso opazili v stopnji celične smrti v starosti kulture povečanega.
Zato omejeni pankreas celičnih kultur izključno povzročila zmanjšanje mitotičnega aktivnost, in brez izgube celic samih. Poleg tega, da je razlog za domnevo, da je zmanjšanje genske aktivnosti je vzrok večine morfološke in biokemične spremembe opazili v celicah starih in odmrlih rastlin.
Na primer, je bilo ugotovljeno, da je opisan na koncu življenjske kulture "degenerativne" celice, ki velik pomen v zgodnjih raziskavah (Hayfiick, Moorhead, 1961- Hayfiick, 1965), ne povzročajo, kot posledica smrti kulture, t. E. Zmanjšajte mitotićnih dejavnost. Ugotovljeno je bilo, da se je veliko tako imenovanih "degenerativnih sprememb" mogoče dobiti v mlado kulturo, zatreti za daljše obdobje delitev celic.
Vse te "degenerativne spremembe," je po več podvojitev prebivalstva izginejo, če ponovno ustvariti pogoje za rast rastlin (Brunk et al., 1973). Povečanje velikost celic pri staranju kultur tudi tako, da upočasni in ustavi celično delitev očitno povzroča. Dejansko, so opazili povečanje velikosti samo v celicah, ki ne sintetizirali DNA (Bowman et al., 1975).
Navedena dejstva kažejo, da je problem staranja celične kulture v bistvu pride do preučevanje vzrokov, mehanizmov in posledic opazili zmanjšanje mitotićnega aktivnosti.
Ta postopek ne nujno povzroči poškodbe in staranje celic in je lahko posledica njihove diferenciacije (Gavrilov, Gavrilova, od leta 1978 Gavrilov, Yaguzhinskii, 1978), ki jo pogosto spremlja zmanjšanje mitotičnega aktivnost (Truman, 1976). Zato je pojem "smrti", "smrt", "staranja" in "pričakovana življenjska doba" celičnih kultur so zelo pogojno, kakor tudi pojem "mlade" in "stare" kulture.
Njihove sklopov uporaba raziskovalcev za preučevanje mehanizmov motenj in poškodb, čeprav še ni bilo dokazano, da sam obstoj teh poškodb in motenj v celični kulturi, in ni bila ugotovljena njihova vloga pri zmanjševanju mitotićno dejavnost.
Zato je priporočljivo, da uporabite druge pogoje, da bi usmerili pozornost raziskovalcev je o tem, kaj se v resnici dogaja, da je. E. Na zmanjšanje mitotićne dejavnosti. so že bile takšne pogoje.
Namesto tega koncepta "življenja", "staranje", "smrt" in "smrt", se predlaga uporaba izraza "replikativnega življenjska doba" celična kultura "replikativnega senescenco", "smrt replikacije" in "replikativnega smrtjo" celične kulture.
Pojem "mlade" in "stare" kulture, je tudi zdaj uporablja s pridevnikom "replikacije". Mladi celice (predelne) imenovane replikativnega ter stari in mrtvi (brez deljenja) - postreplicative.
Na kratko, so stari koncepti včasih dane v narekovajih, s čimer bi poudarili svojo konvencijo (Norwood et al., 1974- Gavrilov, Gavrilova, 1978). Pomembno je, da tudi sedaj je biološka starost kulture določi odstotek celic, ki niso sposobni replikacije sinteze DNA (Cristofalo, Sharf, leta 1973 Bowman et al., 1975).
Trenutno breme ustanovljena naslednje osnovne manifestacijo "staranja" celičnih kultur.
1. Zmanjšana delež celic, primernih za delitev (Merz, Ross, 1969- Cristofalo, Sharf, 1973).
2. Povečanje obdobje mitotičnem cikla. Radioautographic zazna povečanje minimalno obdobje generacije celic v gojenih človeških fibroblastov od 16 do 21-22 ur (Macieira-Coelho et al., 1966). Filmska študija je pokazala povečanje povprečnega časa med oznakama z "staranjem" človeških fibroblastnih kultur od 16,8 do 32 ur (Absher et al., 1974).
3. "staranje" celične kulture poveča inhibitornega učinka gostoto prebivalstva na stopnjo rasti (Macieira-Coelho et al., 1966- Absher et al., 1974- Schneider Mitsui, 1976), tako da je gostota nasičenja pri replikacije starih kulturah približno 2-krat manjši od "mladih" kultur (Schneider, Mitsui, 1976).
Zdieľať na sociálnych sieťach:
Príbuzný
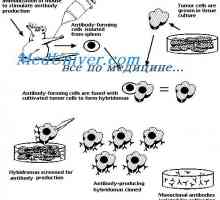 Prevod mRNA. Značilnosti mRNA prevajanja pod sintezo protiteles
Prevod mRNA. Značilnosti mRNA prevajanja pod sintezo protiteles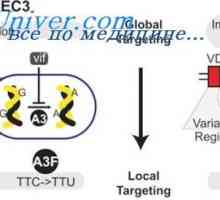 Začetek diferenciacije limfocitov. Shema začetek diferenciacije celic
Začetek diferenciacije limfocitov. Shema začetek diferenciacije celic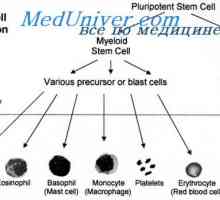 Funkcije in celice. Participativne mehanizme in celično imunost
Funkcije in celice. Participativne mehanizme in celično imunost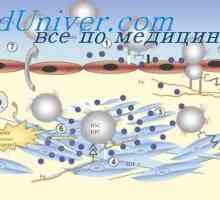 Mehanotsity stromalni limfatične organe. Sevov stromalni mehanotsitov
Mehanotsity stromalni limfatične organe. Sevov stromalni mehanotsitov Vloga in celice v predelavi antigena. Morfologija in celic
Vloga in celice v predelavi antigena. Morfologija in celic Lastnosti in značilnosti T-dušilne člene. Interakcija T-dušilne člene
Lastnosti in značilnosti T-dušilne člene. Interakcija T-dušilne člene Zeleni čaj pomaga varovati telo pred staranjem
Zeleni čaj pomaga varovati telo pred staranjem Citogenetski analiza horionskega villus celice placente.
Citogenetski analiza horionskega villus celice placente.- Hipotalamus-hipofiza ureditev staranja
- Spremembe v delovanju kromatina med staranjem
- Spreminjanje funkcije kromatina med staranjem. prevod spremembe
- Spreminjanje funkcije kromatina med staranjem. S starostjo povezane spremembe sintezo…
- Presnovna vlogo v mehanizmu staranja
- Vloga metabolizma v mehanizmu staranja. presnova ogljikovih hidratov
 Brain staranje
Brain staranje Staranje celic v telesu. Funkcija presnovna podpora celica v starosti
Staranje celic v telesu. Funkcija presnovna podpora celica v starosti Staranje celic v telesu
Staranje celic v telesu- Staranje v celičnih kulturah. delitev Limit celic
- Učinek biološko aktivnih snovi na proces staranja. Vitamini in multivitaminski pripravki
- S starostjo povezane spremembe v genskem naprave celic. spremembe v kromosomih
- Zakoni razvoja staranja