Nevrofiziologija fotorecepcija

Mrežnica izvor in struktura je živčni center, kjer predobdelava pride do svetlobne signale in jih pretvori v živčne impulze.
Nastalo vzbujanje se prenese vzdolž optične poti v korteksa in skorje vizualnih področjih možganov.
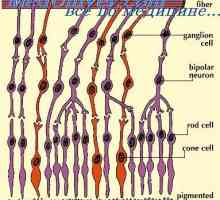
V mrežnici pulz skozi verige 3 glavne vrste živčnih celic:
- fotoreceptorji,
- bipolarne celice,
- celice ganglijev.
Funkcionalna interakcija med njimi poteka skozi amakrinskih in horizontalnih celic v mrežnici.
Nevrofiziološki mehanizmi fotorecepcija zelo kompleksne. Retinala fotoreceptorji temno depolarizirano in končanje kontinuirno izločanje nevrotransmiterjev v sinaptični reži, širine 0.02 mikronov. Nadalje, v temi, temno tok konstantno tkivo tekočina, ki vsebuje natrijeve ione (Na+), Kalij (K+) In kalcij (Ca2+) S ionskih kanalčkov fotoreceptorskih zunanjo membrano segmentov.
Ta stalni fiziološki proces trdi membrana depolarizacija fotoreceptorskih zunanjih odsekov, s čimer se poveča njeno aktivnost.
Svetloba je primerna spodbuda za organ vida. Vzbujanje palic in stožcev mrežnice pod vplivom svetlobe (prevajanje svetlobnih) opremljen in od Bioelektrični zaporedju fotokemičnih procesov, ki se odvijajo v fotoreceptorskih zunanjih odsekih. Fototransdukcijski - ta pretvorba energije absorbira fotonov v kratkoročnem spremembe bioelektrični impedance na plazemske membrane od zunanjega segmenta svetlobnih receptorjev. Pod pride do vpliva svetlobe fotokemične reakcijske vizualne pigmenti in beljakovin del (opsin) beljena en slikovni pigment molekula aktivira veliko število molekul encima fosfodiesteraze in transducina. Po drugi strani pa fosfodiesteraze omogoča kasnejšo hidrolizo (razgradnjo) več deset tisoč guanidinmonofosfata cikličnih molekul (cGMP), s katero je odprta ionskih kanalčkov fotoreceptorskih zunanjo membrano segmentov. Svetloba, povečanje membranski potencial fotoreceptorjem, ga hyperpolarizes, s čimer se zmanjša izločanje nevrotransmiterjev v sinaptični reži. To pa vodi do sprememb v električni potencial v postsinaptičnem nevronu naslednjem poglavju, t. E. V bipolarnih celic. Če je membranski potencial v temi majhna in fotoreceptor je 20-40 mV, kadar je izpostavljen svetlobi fotoreceptorskih zunanjega segmenta membrane in hyperpolarizing membranski potencial je osvetljena svetle poveča za 30-50 mV. Hyperpolarization nadaljuje v celotnem obdobju izpostavljenosti svetlobi.
Tako primarni fotokemične reakcijske vizualne pigmenti na zunanjih segmentih fotoreceptorjih je začetek vizualnega procesa. Kot rezultat, izginja le eno molekulo vidnega pigmenta zaprta več deset tisoč ionskih kanalov fotoreceptorskih membran, ki pojasnjuje izjemno visoko občutljivost človeške oči na zelo šibkih svetlobnih virov, predvsem v temi. V svetlem so zaključene vse ionske kanale membran fotoreceptorjih in nadaljnjo krepitev svetlosti ne vpliva na stanje ionskega kanala in daje dodatno membranski hyperpolarization. Elektrofiziološko manifestacija fotokemičnih procesov v fotoreceptorskih zunanjih segmentov je "A" -wave elektroretinogramu (ERG).
Proces dezintegracije in ponovne sinteze cGMP se uravnava z zelo napredno fiziološki sistem. Sistem uredbe je namenjen zelo hitro povratno odvisnih ionskih kanalov v temnem stanju in s tem tudi temno raven obnavljanje membranski potencial. Mehanizem zunanjega segmenta svetlobe prilagajanje fotoreceptorskih je počasi. Pomembna vloga pri izvajanju teh dveh procesov igrajo kalcijeve ione - klasično znotrajcelično mediatorja. V središču hitro okrevanje temnega ravni potenciala membrane je molekularni mehanizem hitrega odpiranja cGMP-ograjenih ionske kanale. Postopek obnovitve vsebuje "vrzel" iz kanala protein kalcijevih ionov - kalmodulin. Kot rezultat desorpcijo kalmodulin poveča sposobnost vezave ionov kanala molekule cGMP je, kadar je njihova koncentracija po blisk svetlobe v zunanjem segmentu fotoreceptorjem majhna. Kalcij je tudi regulator znotrajcelične aktivnosti nekaterih encimov.
V zunanjem segmentu svetlobnih receptorjev, obstaja tudi encimski mehanizem ponovnega sintezo cGMP. Mehanizem encimska regulacije proteinov, ki sodelujejo, so izrazili antigensko aktivnost. Eden od teh proteinov - arestin. V imunologije, protein znan kot "S-antigen" - topne (topnih) antigen. Arestin - najbolj aktivni uveitogenny bela očesa. Avtoimunske reakcija je pomembno v patogenezo številnih bolezni oči.
Fotoreceptorji so med seboj povezani bioelektrične stike. Njun odnos je zelo selektiven: palice vezani s palicami, storži - s stožci. Glede na barvni vid, povezane s stožci krasnooschuschayuschie krasnooschuschayuschimi občutljivih na zeleno - z zelenooschuschayuschimi, modrih stožcev - z sineoschuschayuschimi.
} {Modul direkt4
Fotoreceptorskih celica sinaptični konča terminalyo, ki ima zapleteno strukturo. Zunanja plast mrežnice sinaptičnih funkcionalno interakcijo tri nevron: fotoreceptor, bipolarna in horizontalne celice. Nevrotransmiterjev v teh sinaps je glutamat. Ko je prenos zagon iz fotoreceptorjem s signalom bipolarne celice pomnožili približno 10-krat. Zato lahko majhne spremembe v fotoreceptorskih potenciala povzroči veliko reakcijo bipolarna.
Obstajata dva načina, v živčni impulzi prenosnih struktur mrežnice. Direktna pot - od svetlobnih receptorjev za bipolarno in z bipolarno do ganglijev celic. Posredno način je povezan s preklapljanjem med svetlobnih receptorjev in bipolarnih horizontalnih celic v mrežnici, in med bipolarnih in ganglijskih celic - amakrinskih celic.
Rod-nevronska pot. V mrežnice, obstaja samo ena vrsta bipolarne celice morfolo tvori spojine s paličasto-oblikovanih svetlobnih receptorjev. Rod-bipolarne zbira vložkov iz palice 15-30 v zunanjem plexiform plasti mrežnice. Bipolarni oblika invaginating odvisnih sinapse med svojimi dendriti in paličastega terminala. Aksoni bipolarne gredo v notranjo plexiform plasti in preneha v mrežnici organih ganglion celic. Vendar pa, kot je razvidno iz študije elektronske mikroskopije, izrastki iz ganglijev celice ne tvorijo neposrednih stikov v aksoni bipolarnih celic. Komunikacija med aksonov in dendritov bipolarne celice ganglijev prek amakrinskih celica.
Tako so drogovi povezani z ganglijskih celice niso neposredno, ampak posredno - amakrinskih celico.
To omogoča, da mrežnica zavedati dveh zelo pomembnih fizioloških procesov:
- razhajanje (sipanje) signala in rod-
- konvergenca (zbirka) signalov iz mnogih palice in paličastega bipolarno njihove sinaptične izhod celice ganglijev.
Amakrinskih celica je povezana tudi z Rod- način stožca, tako da lahko Rod- signali uporabijo pot kolbochkovh bipolarna na ganglijskih celic mrežnice. Razlike in konvergenca v nevronskih paličastih poti vodi do akumulacije in povečano paličastega signala na zelo nizkih ravneh intenzivnosti svetlobe. To omogoča, da se človeški vizualni sistem, da je zelo občutljiv na celo en sam visoki svetlobe.
Nevronov od stožec poti. Prenos signalov iz stožcev v mrežnici bistveno razlikuje od-rod nosi signalni poti. Že v zunanji plexiform plasti sinaps obrazec mrežnice stožci ravni z različnimi vrstami bipolarnih celic, in ne z eno samo vrsto celic, ki paličastega sistema. Od stožec bipolars različnih velikosti dendritičnih procesov, in na tej osnovi razlikujemo dve vrsti bipolarnih zaporedju: Midge (pritlikavec) ali pritlikave miniaturne bipolars, razširjen in razpršenih krupnopolnye bipolars. Od stožca bipolarne celice, drugačne kot paličastega bipolarno po vrsti sinaptičnih stikov.
Tako je od stožca bipolarne razliko ravnih paličastih ki sinapse z dendriti celice ganglijev mrežnice, ne z amakrinskih celico kot vmesnih elementov za prenos signalov. Zato je pot stožca bolj neposredno in manj konvergentno kot Rod-. Ena štetje vsebnika bipolarne konvergira manjše število stožcev od palic eno paličasto-bipolarnih. V zadnjem traktu cone nevronskih vezij zelo majhna količina stožca bipolarno in bipolarni konvergira še ena na ganglijskih celic, ki se nahajajo na fovea. Pri tej vrsti stožec prenosnega signala sredino zagotavlja zelo dobro ločuje (ostrina vida) stožca območju osrednje fovea mrežnice in visokim kontrastom.
Premer osrednjem območju receptivnega polja bipolarnih celic v mrežnici je 100-200 um, kar ustreza razvejenega področja celične DENDRITE bipolarne. Zato je odziv bipolarne celice dovzetni terenu centru, ko svetloba stimulacija povzroča stike z fotoreceptorskih sinaps na DENDRITE bipolarne motnje.
Za visoko ostrino vida od stožca poti iz fovee mrežnice ima dva kanala - na in izklop. V stiku z dvema bipolarno Midge (ob cesti in izven centra) povezave, ena stožec Fovea, ki so v stiku z dvema midge ganglijskih celic (tudi ob cesti in izven centra). V zvezi s tem, je verjel, da je razmerje 1 v območju fovee med bipolarno in debla celice: 1. Vendar pa je zunaj fovee je mogoče povezati več vrst stožec bipolarno enega. Torej, kot je že 3 mm od fovea na bližnji obodu, Midge bipolarne celice povezan z 2-3 stožci. Zato je v vezju stožca na vsaki fovee poti nosi informacije o le eno barvo na sredini receptivnega polja, in iz drugih področij mrežnici stožca večkanalno lahko signalni poti prehaja iz ene vrste stožci ali več vrst, tj. E. Bodite heterochromic. Iz tega razloga, je ostrina vida na mrežnici parafoveal območju bistveno zmanjša.
Od stožec poti pojav tudi dosledno in sočasno kontrast vidnega sistema. Horizontalni celice, povezan preko površine mrežnice v sincicij sinaps zagotavlja funkcijo istočasnega kontrast.
 Histogenezi zarodka mrežnice. Morfologija retinala ploda
Histogenezi zarodka mrežnice. Morfologija retinala ploda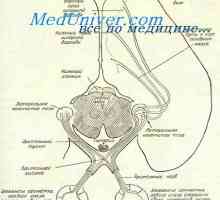 Nastanek živca v oči zarodkov. Poti do vizualnih centrov plod
Nastanek živca v oči zarodkov. Poti do vizualnih centrov plod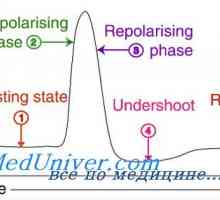 Self-vzbujanje. Mehanizmi lastno vzbujanje celic
Self-vzbujanje. Mehanizmi lastno vzbujanje celic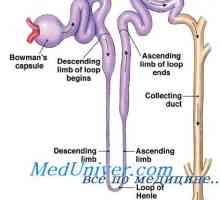 Kalijev sekrecijski primarne ledvične celice. Dejavniki reguliranje izločanja kalija
Kalijev sekrecijski primarne ledvične celice. Dejavniki reguliranje izločanja kalija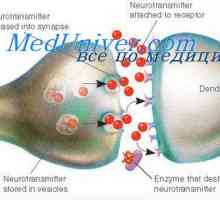 Mediator presinaptičen membrana. postsinaptični membrana
Mediator presinaptičen membrana. postsinaptični membrana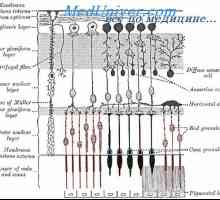 Retin-A. Struktura in anatomija mrežnice
Retin-A. Struktura in anatomija mrežnice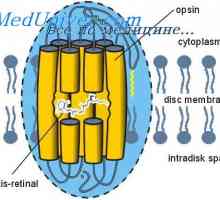 Svetlo in temno prilagoditev. Mehanizmi svetlimi in temnimi prilagajanje
Svetlo in temno prilagoditev. Mehanizmi svetlimi in temnimi prilagajanje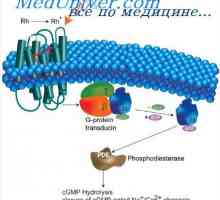 Slepota za posamezne barve. Funkcija mrežnice nevronov
Slepota za posamezne barve. Funkcija mrežnice nevronov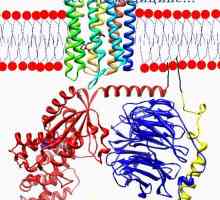 Funkcija horizontalnih celic v mrežnici. bipolarne celice vzbujanje in inhibicijske
Funkcija horizontalnih celic v mrežnici. bipolarne celice vzbujanje in inhibicijske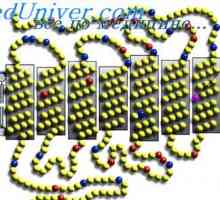 Amakrinskih celic. Mrežnice celice ganglijev
Amakrinskih celic. Mrežnice celice ganglijev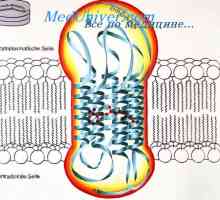 Vizualni pot iz storžkov. Nevrotransmiterji mrežnične nevroni
Vizualni pot iz storžkov. Nevrotransmiterji mrežnične nevroni Vzbujanje celice ganglijev. Vloga lateralne inhibicije v mrežnici
Vzbujanje celice ganglijev. Vloga lateralne inhibicije v mrežnici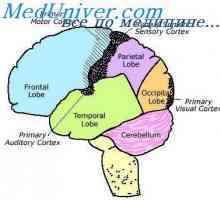 Analiza kontrastov v vizualni podobi. vizualna funkcija korteks
Analiza kontrastov v vizualni podobi. vizualna funkcija korteks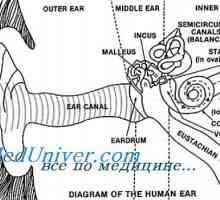 Organ za Corti. Fiziologija in funkcija organa Corti
Organ za Corti. Fiziologija in funkcija organa Corti Nevroni: ključnih izrazov
Nevroni: ključnih izrazov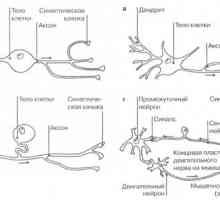 Vrste živčnih celic
Vrste živčnih celic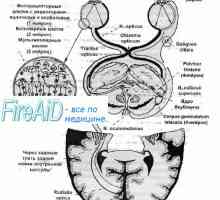 Poti. Pot vizualnega analizatorja. Prevodni pogled na pot.
Poti. Pot vizualnega analizatorja. Prevodni pogled na pot. Patogenetske mehanizmi motnjah funkcije vida pri bolnikih z multiplo sklerozo
Patogenetske mehanizmi motnjah funkcije vida pri bolnikih z multiplo sklerozo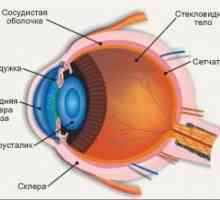 Anatomija in fiziologija mrežnice
Anatomija in fiziologija mrežnice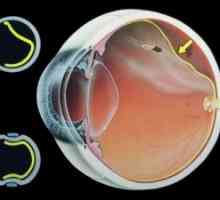 Embriogeneza mrežnica
Embriogeneza mrežnica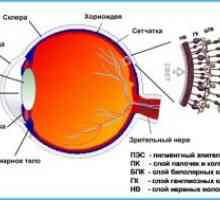 Razvoj senzorične funkcije mrežnice v embriogeneze in zgodnjih letih
Razvoj senzorične funkcije mrežnice v embriogeneze in zgodnjih letih